|
STEFAN KRYNSKI, EUGENIUSZ BECLA, MARIAN MACHEL WEIGL'S METHOD OF INTRARECTAL INOCULATION OF LICE IN PRODUCTION OF TYPHUS VACCINE AND EXPERIMENTAL WORKS WITH RICKETTSIA PROWAZEKI Department of Microbiology, Institute of Pathology, Medical Academy in Gdansk Head: Prof. dr Stefan Krynski |
 METODA WEIGLA DOODBYTNICZEGO ZAKAŻANIA WSZY I PRODUKCJI SZCZEPIONKI PRZECIW DUROWI PLAMISTEMU I PRACACH DOŚWIADCZALNYCH Z RICKETTSIA PROWAZEKI
METODA WEIGLA DOODBYTNICZEGO ZAKAŻANIA WSZY I PRODUKCJI SZCZEPIONKI PRZECIW DUROWI PLAMISTEMU I PRACACH DOŚWIADCZALNYCH Z RICKETTSIA PROWAZEKI
CONTENTS:
- INTRODUCTION
- I. BIOLOGY OF WEIGL'S LABORATORY STRAIN OF BODY LOUSE
- II. THE PROCEDURE IN LABORATORY BREEDING OF LICE
- III. INJECTION OF LICE BY WEIGL'S TECHNIQUE
- IV. BIOLOGY OF BODY LOUSE INFECTED INTRARECTALLY WITH RICKETTSIA PROWAZEKI
- V. THE PROCEDURE IN BREEDING OF LICE INFECTED WITH RICKETTSIA PROWAZEKI
- VI. THE PROCEDURE IN VACCINE PREPARATION
- VII. EPIZOOTIES IN LABORATORY COLONIES OF LICE
- VIII. HARMFUL EFFECT OF LABORATORY LOUSE FEEDING ON HOST ORGANISM
- IX. INFECTIONS OF WORKERS IN LABORATORIES EMPLOYING WEIGL'S METHOD
- REFERENCES
Research works on Rickettsia prowazeki and typhus vaccine production are carried out on Weigl's strain of body louse. These insects are inoculated by intrarectal injections. The following factors affect the course of infection: the strain of Rickettsia prowazeki and the number of its passages through lice, the inoculum size and environmental temperature. High doses of Rickettsia prowazeki have a toxic effect on louse. When the infected insects turn red, they perish. One highly infected louse contains 107LID100 of Rickettsia prowazeki. Epizootics due to cocci. Gram-negative rods, Rickettsia rocha-limae and Rickettsia quintana occur in louse colonies. Feeding of lice can evoke oligo-chromemia in human feeder. Skin reactions and allergy to louse faeces are not uncommon. The workers of laboratories employing Weilg's method can be subject to abortive typhus and trench fever.
INTRODUCTION The first effective vaccine against typhus was made from stomachs of lice infected with Rickettsia prowazeki and the inventor of intrarectal technique of infecting these insects was Rudolf Weigl (1883-1957), an eminent Polish scientist, a professor of University of Lwów, Cracow nd Poznan [29]. A zoologist by education, he was a pupil of Benedykt Dybowski and Jozef Nusbaum-Hilarowicz. He began his investigations on typhus in 1915. Weigl's [60, 61] researches have definitely establish, the pertinence of Rocha-Lima [48] finding that Rickettsia prowazeki is an etiological factor in typhus.
The initial technique of infecting lice was rather complex and highly time-consuming [60, 61]. Microcapillary pipette mounted on a syringe was used for injecting Rickettsia prowazeki suspension. It was only in the years 1938-40 that this method was modified enabling a rapid injection of a big number of lice [15, 19, 20, 49].
Weigl's vaccine developed in its initial version in 1918 was then tested repeatedly on guinea pigs and afterwards on human volunteers, the first 'of them being a technician in Weigl's laboratory, Mrs Rozalia Martynowicz [64, 65, 66, 67]. Next, the evaluation of the vaccine was performed by Radlo [50] and Chodzko [4] in South-Eastern Poland. When the effectiveness and harmlessness of this vaccine had been sufficiently tested it was lent to the Belgian Catholic Mission in China, where it obtained an extremely high assessment [51, 52]. Vaccinations were also carried. out in Africa [33, 34]. Many thousands of people were immunized with Weigl's vaccine during the IInd World-War [42, 43, 68, 69, 70,74].
Weigl's vaccine is admittedly more complicated in preparation and less economic than that of Cox, but in some circumstances this situation may be reversed. Weigl's method has, however, preserved its high value in experimental works and not only in relation to Rickettsia prowazeki but other microorganisms as well [23, 24, 27, 28, 30]. Its modifications have also found application in other arthropods [32, 71]. In view of the fact that Weigl himself had never presented his modified method and his coworkers had contributed to it new ideas it seemed worthwhile to give its detailed description, both from the point of view the vaccine production and experimental works.
I. BIOLOGY OF WEIGL'S LABORATORY STRAIN OF BODY LOUSE All our investigations and the production of Weigl's vaccine in Poland are carried out on lice originated from laboratory colony founded by Weigl in 1918. The insects were taken by him from war camps located at that time in Southern Poland, In 1939 lice from Laboratory for Rickettsiosis Prophylaxis and Research in Addis-Ababa (Ethiopia) were delivered to the Department of Biology at the Medical Faculty of Lwow University and cross-bred there with the Polish strain. Thus the lice employed now are hybrids of the Caucasus race and the African one. So obtained strain has been called by us "Weigl strain".
At 32° the percentage of parous eggs varies from 93 to 96. First individuals appear between the day 3 and 4 and the process is terminated on the day 7 or 8. The peak takes place between the day 5 and 6 of the incubation and from 75 p.c. to 90 p.c. of larvae hatch then. Lowering the environmental temperature down to 28° delays this process by 4 or 5 days without affecting, however, the number of parous eggs. Raising the temperature up to 37° shortens slightly this period, but diminishes the proportion of hatched larvae to 44-76.
 At 32° the first moulting of 95-97 p.c. is observed between the 2d and 3d day, of life. The rest undergoes this process on the 4th day. The second moulting occurs either on the 4th day (40-70 p.c.) or on the 5th day (30-60 p.c.). Sexual maturity of lice is obtained after third moulf which appears between the day 7 and 10. From 30 to 60 p.c. of insects are mature as early as on the 7th day and 90 p.c. on the 8th.
At 32° the first moulting of 95-97 p.c. is observed between the 2d and 3d day, of life. The rest undergoes this process on the 4th day. The second moulting occurs either on the 4th day (40-70 p.c.) or on the 5th day (30-60 p.c.). Sexual maturity of lice is obtained after third moulf which appears between the day 7 and 10. From 30 to 60 p.c. of insects are mature as early as on the 7th day and 90 p.c. on the 8th.
At 28° the first moulting is delayed by 24 hours in comparison with that at 32°, this process occurring, however, during one day. The second moulting takes place on the 6th (90-93 p.c.) and 7th (7-10 p.c.) day. Sexual maturity is reached at 28° between the 10th and 13th day ,it being true for 70-80 p.c. on 11th day and 98th on the 12th. Only single specimens moult between the 12th and 13th day.
3. The weight of larvae and adult insects
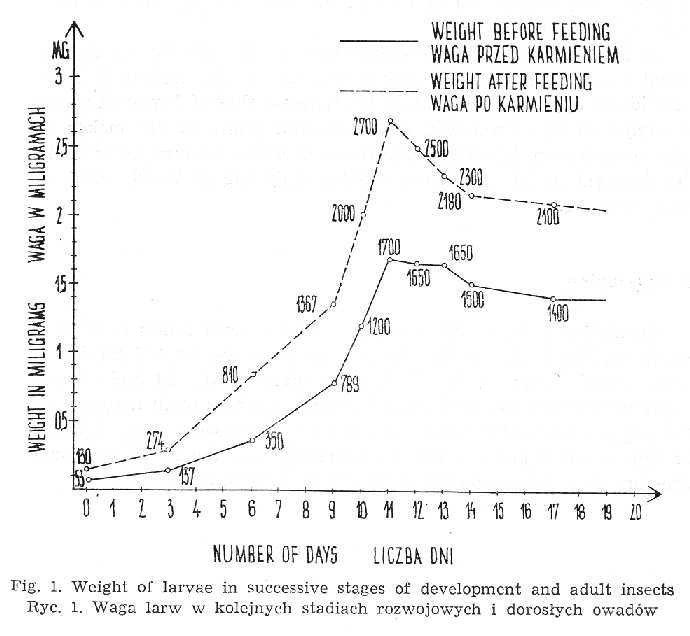
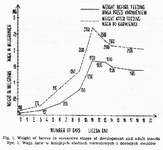
After hatching, the weight of larvae ranges from 0.050 to 0.077 mg. Its increase in successive stages at 32° and after reaching sexual maturity is presented in Fig. 1. Maximal weight of a louse is attained on llth day of life. An unengorged female weighs from 2.0 to 2.5 mg, while the engorged one - from 3.0 to 4.0 mg. In males the figures read respectively: 1.5 and 2.0.
At 28° the weight gain is slower than at 32° with the maximal point being attained on the 14th day o life. At 28° the weight of engorged individuals approximates that at 32°, whereas that of famished specimens is higher at 28°. Prolonged developmental phase at 28° makes larvae take more human blood in order to reach sexual maturity. For their full development at 28° the larvae require 4.352 mg of blood, while at 32° only 3.020 mg.
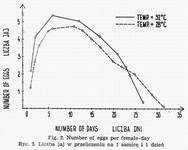
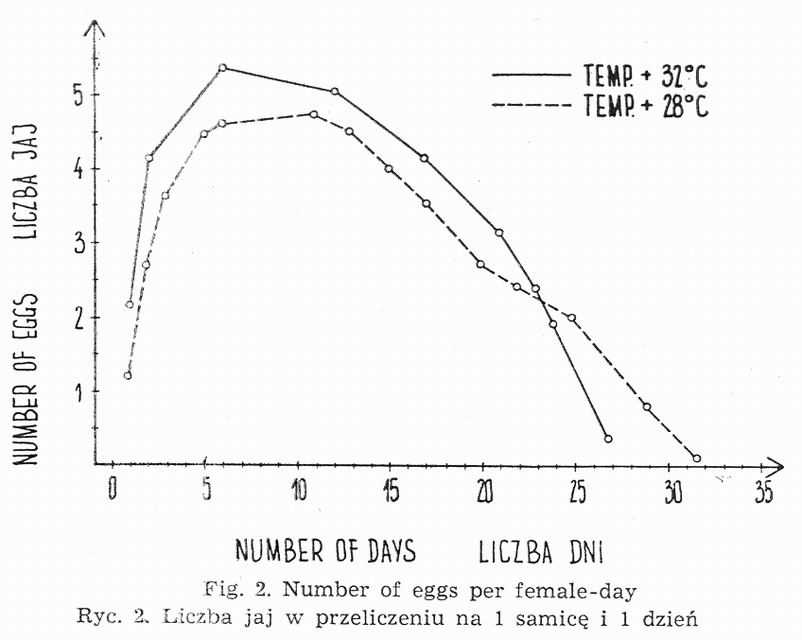
Copulation both at 32° and 28° occurs several hours after the last moulting. Egg-laying at 32° starts already on the 1st and 2nd day following sexual maturity. At 28° it takes place on the 2d and 3d day. W cages holding females and males in equal numbers or in natural proportion, eggs are laid during 23-29 days. Fig. 2 presents mean day number of eggs counted per one female. Generally one female lays during her life-time from 70 to 100 eggs at 32° and 50 to 90 at 38°.
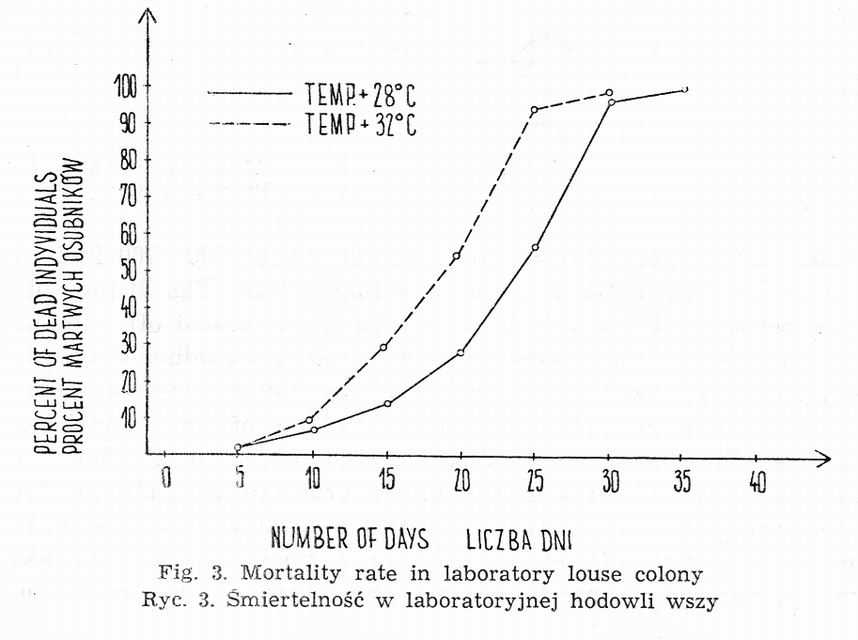
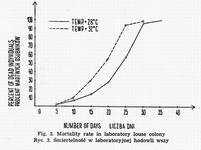
At 32° maximal survival time of males in a given population amounts, on average, to 26.4 days, while in females it equals 28.7 days. At 28° there are no apparent differences associated with the sex. Total depopulation is observable between the 34th and 36th day (Fig. 3).
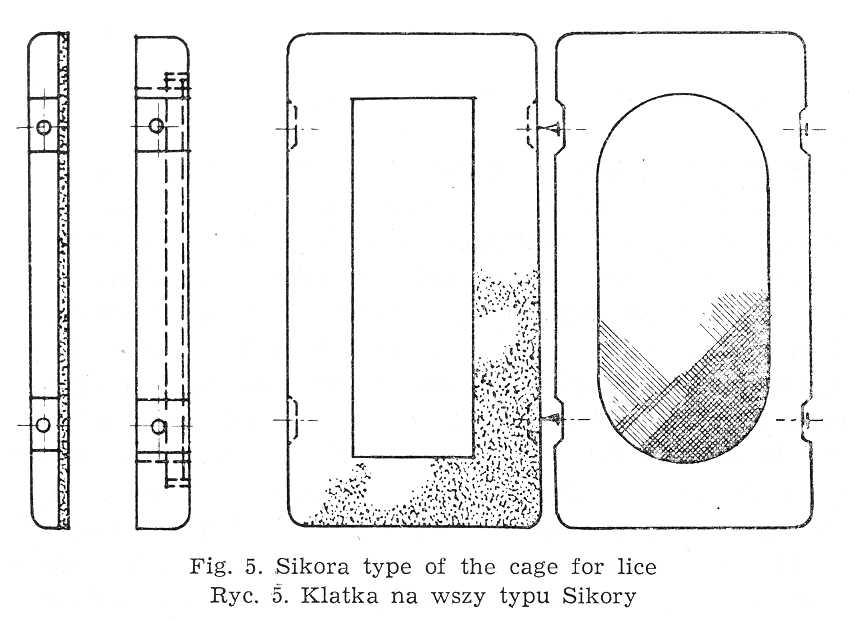
II. THE PROCEDURE IN LABORATORY BREEDING OF LICE  A scrap of cloth with louse eggs is placed inside a tube, with the narrowing at is lower part (Fig. 4) plugged with cottonwool. At 32° larvae start to hatch after 3 days of incubation and subsequently drop down to the bottom of the tube. Spilled-out on Petri dishes, newly hatched larvae are collected on a strip of cloth or wool and next put into sterile cages provided with a net made of bolting-doth. About 600-700 individuals are placed m each of them. In cages of Sikora type (Fig. 5) the point of contact between the lid and frame of the net is sealed off (with paraffin or collodium. Three cages each are put into cardboard boxes, net downwards. The boxes are provided at the bottom with strips 5 mm broad and 3 mm thick. It safeguards the access of air to the inside of each cage. The louse faeces collects at the bottom of the box and is examined every day on agar medium in order to ascertain the asepsis of lice. Larvae likewise adult insects are fed on human volunteers, mostly on legs, once daily. Hands, leg skin and a rubber belt used for keeping them fast must be disinfected with 60 p.c. alcohol.
A scrap of cloth with louse eggs is placed inside a tube, with the narrowing at is lower part (Fig. 4) plugged with cottonwool. At 32° larvae start to hatch after 3 days of incubation and subsequently drop down to the bottom of the tube. Spilled-out on Petri dishes, newly hatched larvae are collected on a strip of cloth or wool and next put into sterile cages provided with a net made of bolting-doth. About 600-700 individuals are placed m each of them. In cages of Sikora type (Fig. 5) the point of contact between the lid and frame of the net is sealed off (with paraffin or collodium. Three cages each are put into cardboard boxes, net downwards. The boxes are provided at the bottom with strips 5 mm broad and 3 mm thick. It safeguards the access of air to the inside of each cage. The louse faeces collects at the bottom of the box and is examined every day on agar medium in order to ascertain the asepsis of lice. Larvae likewise adult insects are fed on human volunteers, mostly on legs, once daily. Hands, leg skin and a rubber belt used for keeping them fast must be disinfected with 60 p.c. alcohol.
After drying of the skin, a dry belt is fastened round the leg and cages with nets towards the skin are inserted beneath it. The duration o the feeding usually amounts to 30 minutes. In persons acting as feeders for many years it I must be sometimes prolonged by next 15 minutes.
 After 5 or 6 days larvae should be transferred into new cages in view of the fact that faeces is partially retained in the meshes of the net, hampering the access of air and blood sucking. Vapour collects inside the cages causing the louse body to be wet and facilitating the adhesion of faeces to it. Asphyxia may result in these conditions. It also makes bacterial infections easier. On the 7th or 8th day, the larvae should be introduced into new cages. This time not more than 300 specimens are placed into one cage. In more populated cages, applied previously in Weigl's laboratory, bacterial epizootics occurred fairly frequently. Indusiae are removed from cages on the 10th day and the lice, in case of need, are transferred to a new cage. After 12 days all lice are mature and large enough to be infected with Rickettsia prowazeki. If the lice are earmarked for reproduction they are kept for next 6 or 8 days, the cages being changed after 4 days. The cloths with eggs are put into the hatching tubes. More economic than reproductive breeding is making use of eggs of infected lice. It presents no risk, as Rickettsia prowazeki does not cause any transovarial infections in lice.
After 5 or 6 days larvae should be transferred into new cages in view of the fact that faeces is partially retained in the meshes of the net, hampering the access of air and blood sucking. Vapour collects inside the cages causing the louse body to be wet and facilitating the adhesion of faeces to it. Asphyxia may result in these conditions. It also makes bacterial infections easier. On the 7th or 8th day, the larvae should be introduced into new cages. This time not more than 300 specimens are placed into one cage. In more populated cages, applied previously in Weigl's laboratory, bacterial epizootics occurred fairly frequently. Indusiae are removed from cages on the 10th day and the lice, in case of need, are transferred to a new cage. After 12 days all lice are mature and large enough to be infected with Rickettsia prowazeki. If the lice are earmarked for reproduction they are kept for next 6 or 8 days, the cages being changed after 4 days. The cloths with eggs are put into the hatching tubes. More economic than reproductive breeding is making use of eggs of infected lice. It presents no risk, as Rickettsia prowazeki does not cause any transovarial infections in lice.
III. INJECTION OF LICE BY WEIGL'S TECHNIQUE 1. Preparation of Rickettsia prowazeki suspension
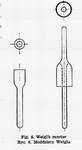
 Preparation of Rickettsia prowazeki suspension [19] varies in dependence on its destination. In vaccine production one suspension should be made centrally, as it facilitates future control. "A higher number of infected louse stomachs is then used and suitable dilutions of the suspension are later performed. Best results are obtained employing the concentration corresponding to 0.1 - 1 stomach in 1 ml (106-107 LID100). The faeces of lice earmarked for suspension must be inseminated after last feeding on agar plate, blood agar plate and broth with glucose. Live and bright red lice are taken for the suspension six hours after feeding. In dead insects the proportion of living rickettsiae may be lower, having adverse effects on infectivity of Rickettsia prowazeki suspension. Lice are dissected in a drop of saline and then homogenized in Weigl's mortar (Fig. 6). Saline or preferably buffer N-7 solution [2, 3] are added next in required quantity. K-7 buffer solution more advantageous to Rickettsia prowazeki cannot be applied on account of its harmful effect on louse.
The addition of human serum [55] and particularly of skim milk [1, 21] increases infectivity and stability of a suspension. Bidistilled water should be used for preparation of the suspension, as the presence of copper salts poses a great threat [18].
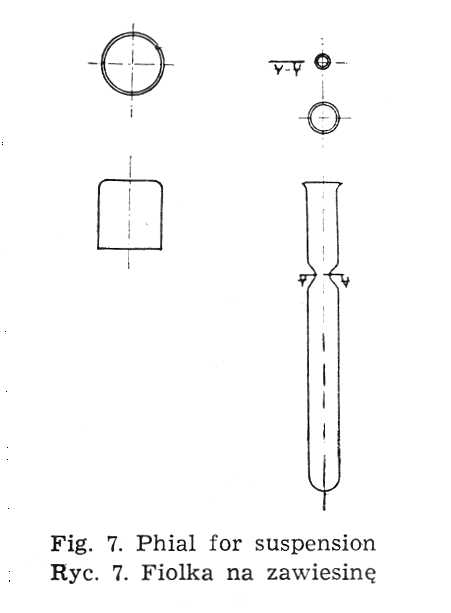
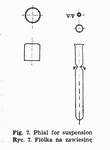
The suspension poured out into phials (Fig. 7) is stored in the refrigerator. The storage should be as brief as possible because even in such conditions it may be disadvantageous.
Preparation of Rickettsia prowazeki suspension [19] varies in dependence on its destination. In vaccine production one suspension should be made centrally, as it facilitates future control. "A higher number of infected louse stomachs is then used and suitable dilutions of the suspension are later performed. Best results are obtained employing the concentration corresponding to 0.1 - 1 stomach in 1 ml (106-107 LID100). The faeces of lice earmarked for suspension must be inseminated after last feeding on agar plate, blood agar plate and broth with glucose. Live and bright red lice are taken for the suspension six hours after feeding. In dead insects the proportion of living rickettsiae may be lower, having adverse effects on infectivity of Rickettsia prowazeki suspension. Lice are dissected in a drop of saline and then homogenized in Weigl's mortar (Fig. 6). Saline or preferably buffer N-7 solution [2, 3] are added next in required quantity. K-7 buffer solution more advantageous to Rickettsia prowazeki cannot be applied on account of its harmful effect on louse.
The addition of human serum [55] and particularly of skim milk [1, 21] increases infectivity and stability of a suspension. Bidistilled water should be used for preparation of the suspension, as the presence of copper salts poses a great threat [18].
The suspension poured out into phials (Fig. 7) is stored in the refrigerator. The storage should be as brief as possible because even in such conditions it may be disadvantageous.
In the culture of strains the suspensions are made from single stomachs, Histological examination [76] must be carried out periodically with the aim of excluding the presence of Rickettsia quintana. It becomes of particular importance during the laboratory epidemics of trench fever, Woyciechowska [75] recommends an expeditious making of histological preparation from half a stomach and preparing a suspension from the other half only after establishing there the absence of extra--cellular rickettsiae.
In experimental works Rickettsia prowazeki suspensions are made from 10 stomachs in 10 ml of buffer solution (107 LID100).
2. Technique of intrarectal inoculation of lice
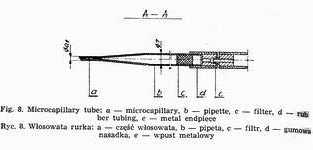
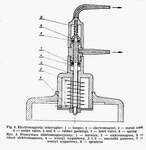
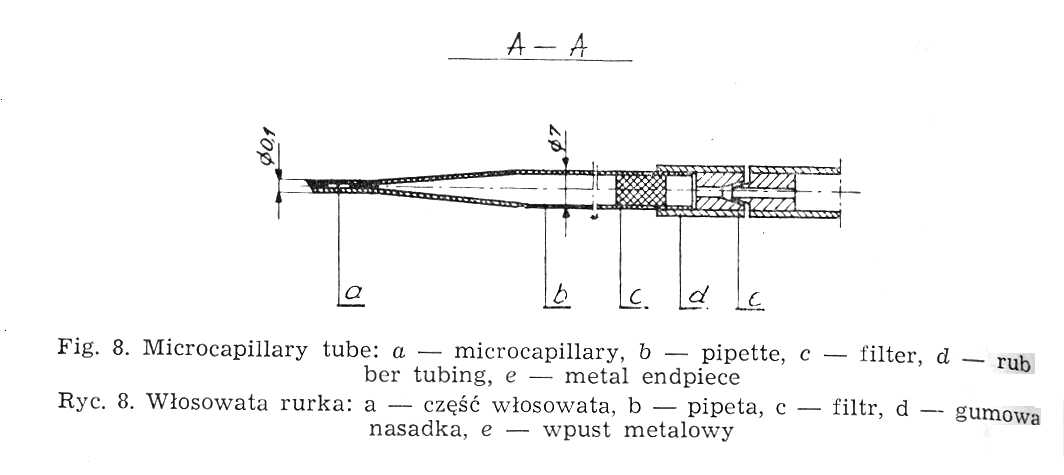
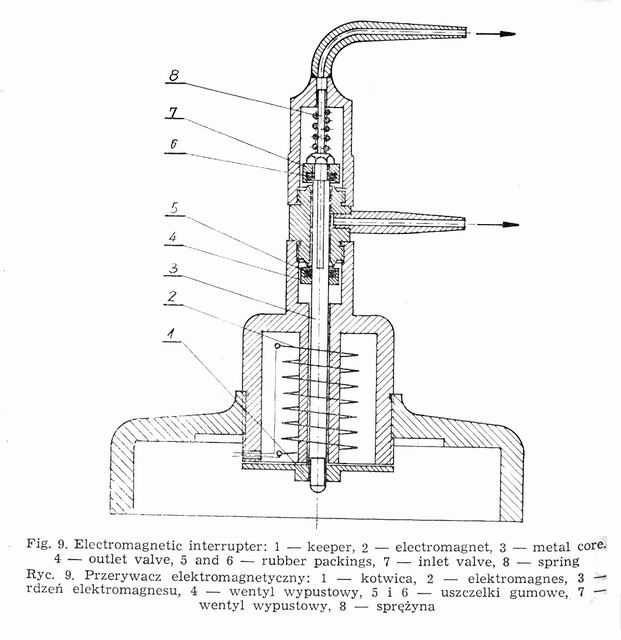
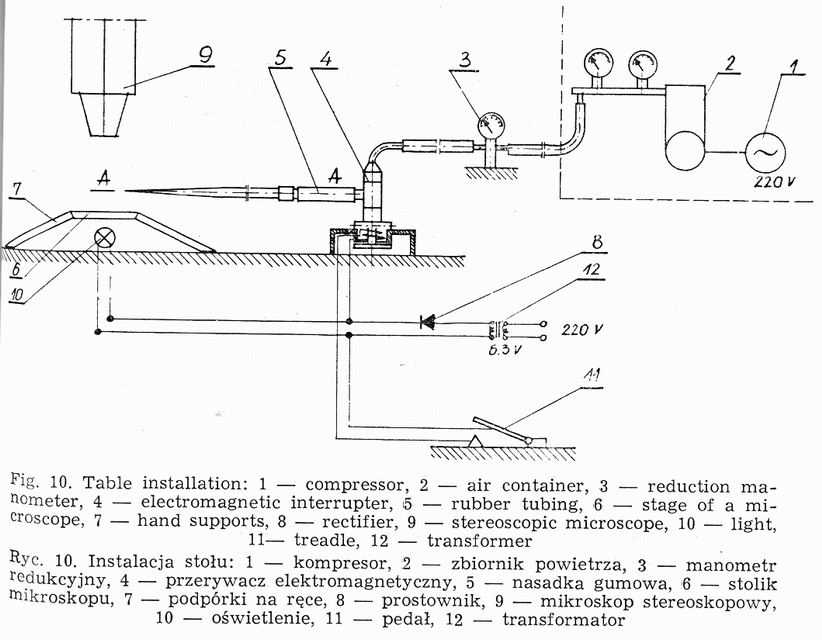
Weigl's methods of louse infection consists in intrarectal introduction of Rickettsia prowazeki by means of a microcapillary (Fig. 8) under Pressure [20, 49]. The air stream flows from an air container through reduction manometer, in which the pressure is reduced to 2.0-2.5 atm. and through an electromagnetic interrupter (Fig. 9) into the capillary tube. The interrupter acts on the principle of electromagnet, to which current is brought through a rectifier, changing it from intermittent +Q a constant one and reducing the tension from 220V to 6V.
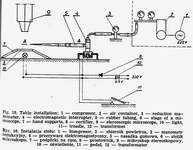 Current circuit is put on by a treadle, the pressure of which causes the lifting of a metal core inside the interrupter. The core is provided in its upper part with rubber packing (inlet valve), constituting a barrier for the air flowing from the container. The lifted core enables a free passage of air through the outlet valve of the interrupter into the rubber tube and finally into the capillary, forcing the suspension out of it (Fig. 10)
Current circuit is put on by a treadle, the pressure of which causes the lifting of a metal core inside the interrupter. The core is provided in its upper part with rubber packing (inlet valve), constituting a barrier for the air flowing from the container. The lifted core enables a free passage of air through the outlet valve of the interrupter into the rubber tube and finally into the capillary, forcing the suspension out of it (Fig. 10)
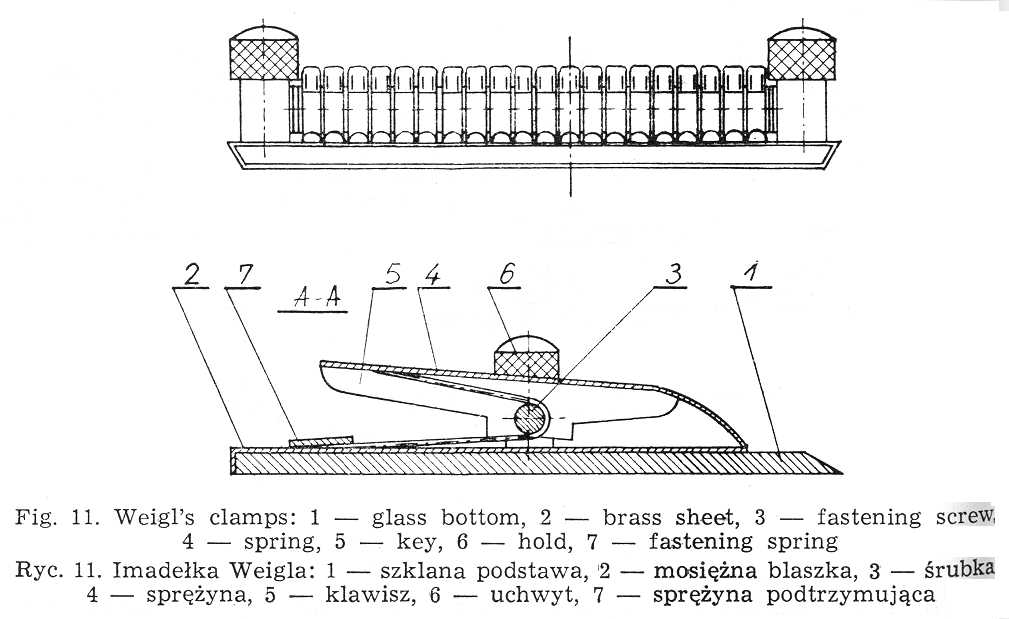
Weigl's micro capillary is made from a Pasteur pipette, the ending of which is extended over a microburner into a capillary. Under the magnification of 32 times the microcapillary is broken off aslant at the angle of 35°-40° in the place where its diameter amounts to 0.1-0.05 mm. The sharp edges are melted off by being moved quickly and repeatedly over a microburner, the degree of melteness being checked under the magnification to ensure the best possible smoothness. A correct made microcapillary should safeguard the lice from getting damaged during the intrarectal injection. The upper part of the pipette, 1.5-2.0 cm from the end, must be narrowed for the cottonwool to be placed there as an air filter. Pasteur pipettes, the micro capillaries are mad from, should be sterilized at 100°. Higher temperature would make the glass brittle, which would hamper precise breaking off of the microcapillary tips. A thick-walled rubber tube with a metal endpiece is tightly placed on the other end of the pipette. If a liquid is to be drawn the nipple of a syringe is inserted into this endpiece. Lice destined for inoculation are immobilized by special Weigl's clamps (Fig. 11), constructed in the following way. A brass sheet is fixed to a glass base. Along its middle line run twenty half circular keys cut out from underneath and mounted each on a spring to safeguard proper elasticity permitting the immobilization of arthropods by pressure. The clamps are sterilized with burning cotton plug moistened with alcohol.
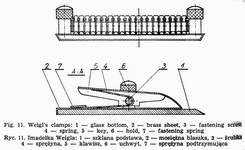 Lice are placed in the lower part of a Petri dish on a black cloth while its upper part is turned over with bottom upwards to accommodate the clamp. It is held with the thumb and third finger of the left hand and the respective key is pressed by the index finger. With the right hand using pincers the lice are caught by the end of the abdominal part and placed successively under the keys. Of great importance here is phototaxic phenomenon causing the movement of lice away from the source of light towards the clamp.
Lice are placed in the lower part of a Petri dish on a black cloth while its upper part is turned over with bottom upwards to accommodate the clamp. It is held with the thumb and third finger of the left hand and the respective key is pressed by the index finger. With the right hand using pincers the lice are caught by the end of the abdominal part and placed successively under the keys. Of great importance here is phototaxic phenomenon causing the movement of lice away from the source of light towards the clamp.
The inoculation of lice is accomplished under stereoscopic microscope at the magnification of 32 times. The microscope should be provided with hand supports. The clamp with lice is placed on the glass table of the microscope on the left side of the field of vision, whereas the micro-capillary with suspension is put horizontally at a suitable height on the right side. This microcapillary is joined by rubber tube with the electromagnetic interrupter. The ending of microcapillary is introduced into ampulla recti of the louse and then the treadle, closing the current circuit, is pressed shortly and the compressed air forces the suspension into the intestinal lumen.
The injected lice are transferred over the open Petri dish and the insects are spilled on it by simultaneous pressing of all keys. Next the individuals (maximum 400) are put with a piece of cloth into a cage.
A team of two persons, one placing the lice in the clamps, the other injecting the insects is most convenient and can inoculate as many as 1500 specimens per hour.
In cases when a liquid of higher density, such as difibrinated blood or the suspension of homogenized animal organ, is to be injected, a microcapillary of broader lumen (0.3-0.5 mm of diameter) should be employed. In some investigations on the effect of chemotherapeutic substances in vitro, the microcapillaries must be coated with a protective layer of India ink in order to avoid the photodynamic action of light.
IV. BIOLOGY OF BODY LOUSE INFECTED INTRARECTALLY WITH RICKETTSIA PROWAZEKI The course of infection of louse inoculated intrarectally with Rickettsia prowazeki depends on strain, inoculum size, environmental temperature, variation of natural susceptibility of a given louse colony and its particular members [9, 16, 17, 22].
1. The course of acute form, of infection
A. Typical acute form of infection
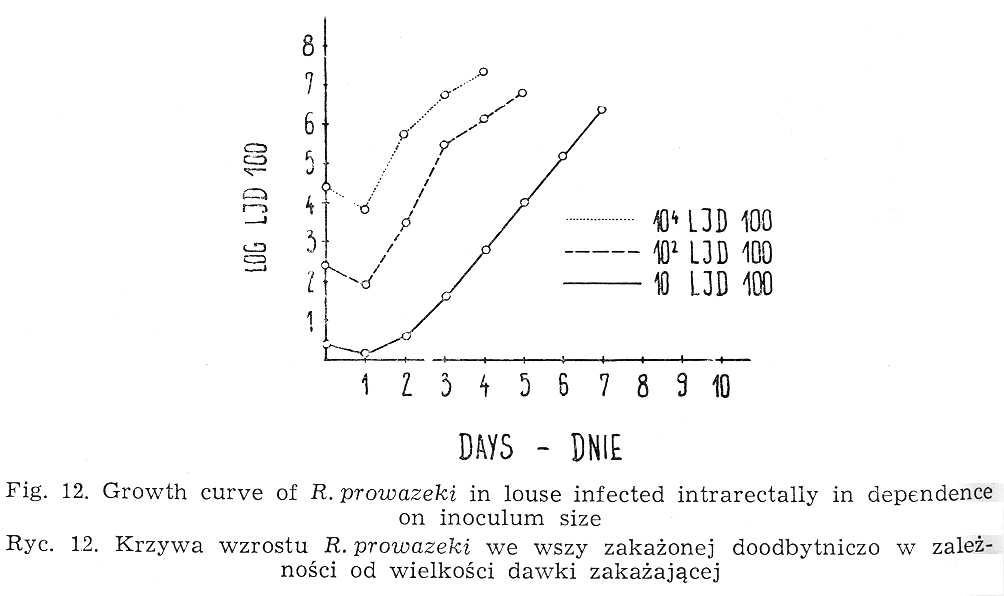
As Weigl [60, 61, 62, 63] has proved, Rickettsia prowazeki inoculate intrarectally begins to enter epithelial cells of louse stomach already after the lapse of 5 minutes. At 32° the process of invasion is over at about the 3d hour of incubation. Only a part of injected rickettsiae penetrate into the cytoplasm of intestinal epithelium cells. A considerable majority - up to 90 p.c. is eliminated with faeces. At 32° the law phase usually terminates between the 18th and 24th hour of incubation {Fig. 12). The duration of the logarithmic phase varies even within the same population of insects. It is also related to the strain and, of course, to the inoculum size [26].
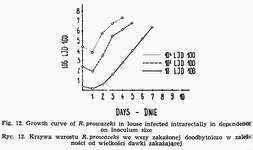
At the beginning of logarithmic phase multiplying ricikettsiae agglomerate in clumps, showing a great variety in morphology. In further stages these clumps become larger and larger to finally fill up profusely the cytoplasm causing the extension of parasitized cells followed by their rupture and, sometimes, detachment from the basal membrane. So, according to Weigl [60, 61], terminates the first period of Rickettsia prowazeki infection in louse. Rickettsiae released into the intestinal lumen start to attack healthy epithelial cells and the second period of louse infection is thus initiated. As Weigl cytological investigations have shown, the growth of Rickettsia prowazeki is considerably less abundant here, because the louse perishes sooner than the logarithmic phase is fully developed. At this stage the evidence of rickettsiae multiplication,is not demonstrable using quantitative methods, because parallel to their growth in newly infected cells there ensues the process of dying out of the bacteria being in the intestinal lumen. The infection degree of lice is the higher the more cells have been invaded immediately after the injection of Rickettsia prowazeki suspension.
The damage of stomach epithelium of louse causes permeation of undigested hemoglobin and even whole erythrocytes into hemocoelom and lymph of insects, making them red. Soon afterwards the infected individuals die.
B. Paralytic form of infection
Louse infected with Rickettsia prowazeki sometimes happens not to become red. One of the causes may be the slow rate of Rickettsia prowazeki growth, which occurs particularly in first passages through lice or at lower temperatures. It is due to the fact that physiologic death precedes the damage of stomach wall (chronic form of infection). Much greater role in vaccine production and experimental works is played by lice which fail to redden due to the lack of hemoglobin in alimentary tract. Paralysis of oesophagus muscles may result from toxic action of Rickettsia prowazeki. The insect pierces the skin of the host, but is unable to suck the blood. The content of the alimentary tract vanishes entirely, the louse assuming white-yellowish hue. Such lice were referred to in Weigl's laboratory as "paper lice". They are very highly infected. If not removed earlier from cages they dry up, which leads to material losses in vaccine production and reflects adversely on the interpretation of the results. Therefore, while checking the cages with lice 3 to 6 h after last feeding, such specimens should be removed. In experimental works they are regarded as red lice.
2. Evaluation of infectivity of louse infected with Rickettsia prowazeki
A. The total lice infecting dose (LID100)
Typhus rickettsiae can be measured by virtue of their capacity to cause infection in louse. Titration is accomplished by the administration of serial 1:10, 1:102, 1:103, 1:104, 1:105, 1:106 and 1:107 dilutions of suspension containing one infected louse stomach in 1 ml of N-7 buffer solution. Each of these dilutions is injected into from 50 to 100 lice. Administration of rickettsiae in the above mentioned doses permits to determine the concentration causing 100 per cent infection among inoculated individuals as well as that producing no infection at all. The lowest dose giving 100 per cent incidence of positive inoculation is designated as a total lice infecting dose - LID100 [25, 26, 31]. One hundred per cent end-point is preferred in experiments on lice in view of the fact that their death may be caused not only by Rickettsia prowazeki infection, but also by other pathological agents, such as mechanical and thermal damages, hindering greatly the determination of the 50 per cent end-point. In border-line cases the proper interpretation of the presence of infection in the damaged insects may be troublesome and even sometimes impossible due to their complete desiccation. Considering that all red lice are infected, those which have failed to become red should be examined microscopically. If the presence of rickettsiae is established in all louse intestinal smears, it can be safely assumed that the unexamined damaged lice are also infected. When, however, some of the insects which have not reddened are germ free, it is not a 100 per cent infection and in such situation the damaged lice are of no interest to us. While determining the 50 per cent end-point we must have precise knowledge of the presence of infection in each louse.
The term "Red lice index" [9, 19, 25, 26] has been introduced while considering the following two parameters:
1.the time lapsing from inoculation of typhus rickettsiae suspension to the liquidation of a given infected population, and
2. the percentage of red individuals received at that time. It is calculated by dividing the percentage of red lice by the number of days.Table I shows the relationship between LID100 and red lice index at 32° using Rickettsia prowazeki strains multipassaged through lice. Each red lice index being applied should be standardized for each strain by means of LID100 determination. While dealing with strains only recently passaged through lice, it must be periodically checked until the stabilization of its virulence for lice.
 Dilutions of control suspension are performed in experimental works in order to determine its LID100 and then the indices of red lice i11 particular dilutions are compared with those of experimental populations.
Dilutions of control suspension are performed in experimental works in order to determine its LID100 and then the indices of red lice i11 particular dilutions are compared with those of experimental populations.
3. Microscopic examination of infected lice
Lice beginning to turn red 'are dissected in a drop of saline, then the stomachs are transferred using a pipette to a fixative of the following composition: 1 part of undiluted formol and 3 parts of 4 p.c. potassium bichromate. About 2 ml of this liquid are sufficient for 10 stomachs. A vessel containing the fixative with louse stomachs should be tightly closed. The time of fixation amounts to 10 min. After fixation, the stomachs must be washed repeatedly in distilled water and placed for at least 10 min in potassium bichromate. The louse stomachs can be kept in the latter even for several months without any deleterious effect on the quality of future histological preparations. In the next stage the stomachs are rinsed with distilled water and dehydrated in alcohols starting with 70 p.c. ethyl alcohol. The material under study is transferred from xylene II to paraffin solution in xylene for 10 min at 60° with subsequent embedding of groups of 3-10 stomachs in paraffin with melting point between 56°-60°. Paraffin sections of 2-3 micr. thickness are mounted on a cover slip. After removing the paraffin, the preparation is put into Giemsa dye made up in the following way; 3.8 g of dry substance is added to 250 ml of glycerol and 250 g of methanol, 4 drops are taken from it and are added to 1 ml of distilled water, then it is alkalized by 0.2 p.c. potassium carbonate to pH 7.2 - 7.4. The time of staining is from 1.5 to 2 h at room temperature. It is differentiated in absolute alcohol and absolute alcohol with xylene aa. The preparation is next embedded in Canada balsam. Rickettsia prowazeki is stained metachromatically and intensively into dark violet color and Rickettsia quintana into cherry-violet [76].
Stomach smears can be stained using all methods employed for rickettsiae, but the best results are obtained with the Giemsa one. Preparation is fixed in mathanol and stained with Giemsa dye of pH 7.0 -7.4 at room temperature during 1.5-2 h. It is differentiated in absolute alcohol with xylene aa [76]. Negative staining is applied to achieve rapid results. Cyanochine [48] or waterblue seem to be most suitable for this purpose.
4. Factors influencing the development of infection and time period red lice occurrence
A. The role of Rickettsia prowazeki strain and the number of its passages
During the World War II we examined a great number of strains deriving from the epidemics in Poland and Ukraine. We found the existence of some differences among them in respect of their virulence for lice. Taking into consideration the time of infection development and toxicity for louse we could distinguish the following 3 groups of strains:
1. Rapid development of infection and high toxicity for louse. Such strains were isolated from ill people during epidemics in Lublin district in the years 1942-1945 and from German soldiers in the years 1942-1943. This group of strains appeared to be best suited for vaccine production and experimental works.
2. Slow development of infection (due perhaps to prolonged lag phase) and very high toxicity not only for lice but also for human beings. Representatives of this group derived from German soldiers returning from Cuban war front.
3. Slow development and low toxicity for louse. Such strains were found during epidemics in Lwow ghetto. This group of strains was very difficult to maintain in louse culture and useless in typhus vaccine production. They perished very often after a few passages through lice.
The first five to ten passages through lice, independently of the above mentioned group attachment, are of irregular character in respect of the time of louse reddening. The same phenomenon is also observed after passaging through animals or yolk sack of chick embryo. This irregularity makes the use of strains in first passages disadvantageous for vaccine production and experimental works and causes their avoidance. We employ strains (Belzyce, Bog., Wolyn and Tomaszow) cultivated since 1945 exclusively on lice.
a. Normal and diluted suspensions
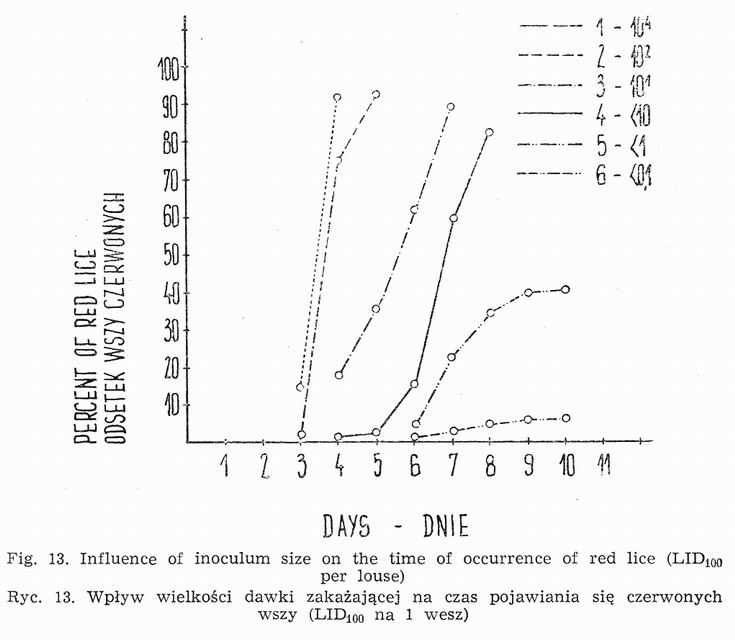
The inoculum size affects the course of logarithmic phase of rickettsia growth (Fig. 12). It lasts the longer the smaller the infecting dose. No differences in the duration of lag phase can be established. In view of the fact that the penetration of hemoglobin into louse hemocoelom takes place only after the termination of logarithmic phase, the occurrence of red individuals is closely connected with its duration. Using the inoculum lO3-4LIDl00 per louse at 32° and strains multipassaged through lice, first red specimens can be observed between 48th and 72d h of incubation (Fig. 13).
 Their percentage is rather low, varying from 5 to 20. About 50 p.c. of the lice in question are strongly infected (l07LID100), about 30 p.c. - moderately (l05LID100) and about 20 p.c. - weakly (105LID100). Most insects become red between 72d and 96th h. Among them strongly infected specimens constitute an overwhelming majority, as much as 80 p.c. There is only 5 p.c. of those weakly infected. A small proportion, not exceeding 20 p.c., turn red later on.
Their percentage is rather low, varying from 5 to 20. About 50 p.c. of the lice in question are strongly infected (l07LID100), about 30 p.c. - moderately (l05LID100) and about 20 p.c. - weakly (105LID100). Most insects become red between 72d and 96th h. Among them strongly infected specimens constitute an overwhelming majority, as much as 80 p.c. There is only 5 p.c. of those weakly infected. A small proportion, not exceeding 20 p.c., turn red later on.
Reduction of the inoculum to 102LID100 per louse decreases the frequency of red lice appearance between 72d and 96th h of incubation (Fig. 13). They are weakly infected. In most insects the morbid process ends on the 4th or 5th day. Likewise at higher doses a certain proportion of individuals survives longer. The percentage of strongly infected lice is lower (about 10). Employing inocula from 1 to 10 LID100 per louse the morbid process is not only prolonged, but the final degree of infection it lower, not exceeding 106LID100. It is due to a smaller number of epithelial cells of louse stomach being attacked during the first period of infection. Greater individual differences become also noticeable, thanks to which the occurrence of red lice is extended over a longer period of time.
Further dilutions of rickettsia suspension cause that not all lice get infected. These insects which display the presence of rickettsiae turn red between the 6th and 10th day after the inoculation (Fig. 13).
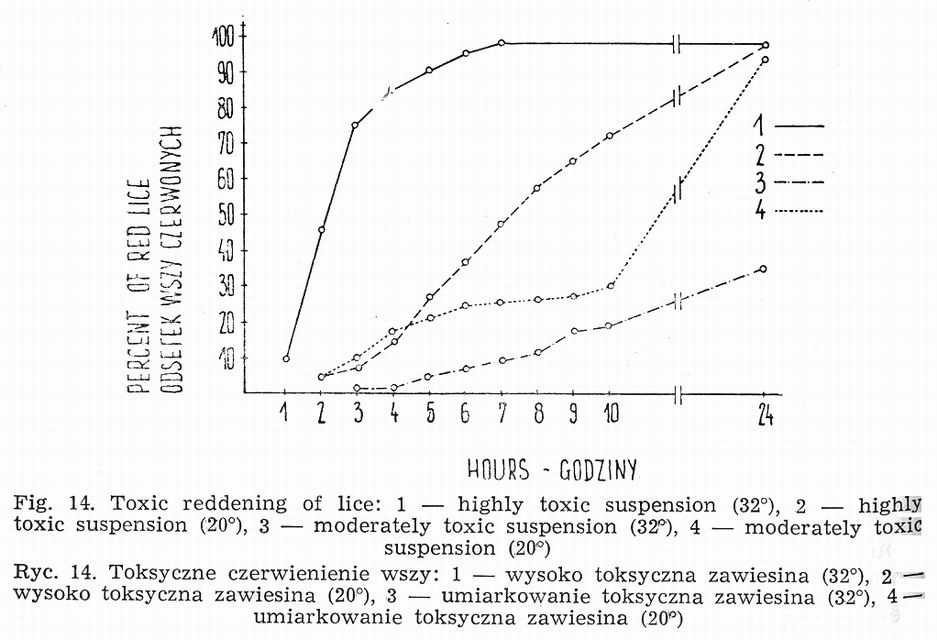
b. Toxic effect of Rickettsia prowazeki in louse
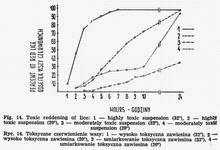 A suspension made of 10 or more infected louse stomachs in 1 ml (1O5LIDl00 per louse) can cause the reddening and death of lice within a few hours (Fig. 14), still before the beginning of the logarithmic phase of growth [12, 14, 17, 22, 31]. The presence of single rickettsiae and the
destruction of epithelial cells of red louse stomach can be established in histological examination. This toxic form of Rickettsia prowazeki infection occurs not in all specimens of a given population. The proportion of lice turning red in the first 24 h after rickettsiae injection depends on the strain and its virulence for lice, the concentration of a suspension, the environmental temperature, the susceptibility of a given population as a whole as well as of single individuals. Employing more concentrated suspensions and highly virulent strains stronger effect is obtained at higher temperature (32°-37°), while at weaker suspensions and less virulent strains - at the lower ones (20°-28°). Toxic action of Rickettsia prowazeki is observed also at 4°. At this temperature red lice appear between the 12th and 48th h of incubation.
A suspension made of 10 or more infected louse stomachs in 1 ml (1O5LIDl00 per louse) can cause the reddening and death of lice within a few hours (Fig. 14), still before the beginning of the logarithmic phase of growth [12, 14, 17, 22, 31]. The presence of single rickettsiae and the
destruction of epithelial cells of red louse stomach can be established in histological examination. This toxic form of Rickettsia prowazeki infection occurs not in all specimens of a given population. The proportion of lice turning red in the first 24 h after rickettsiae injection depends on the strain and its virulence for lice, the concentration of a suspension, the environmental temperature, the susceptibility of a given population as a whole as well as of single individuals. Employing more concentrated suspensions and highly virulent strains stronger effect is obtained at higher temperature (32°-37°), while at weaker suspensions and less virulent strains - at the lower ones (20°-28°). Toxic action of Rickettsia prowazeki is observed also at 4°. At this temperature red lice appear between the 12th and 48th h of incubation.
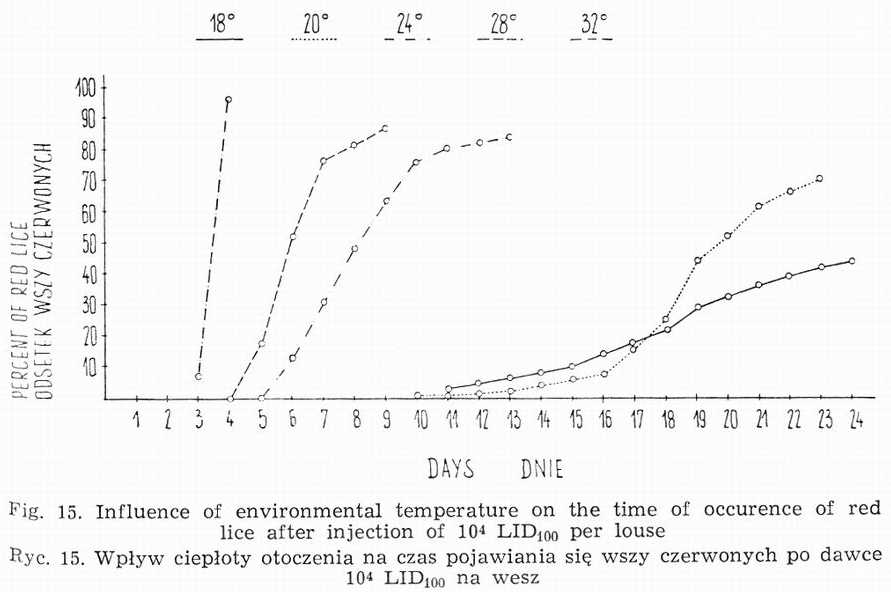
c. Influence of environmental temperature
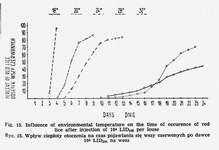 The environmental temperature of 32° is optimal both for lice and rickettsiae. Its rise to 37° does not essentially accelerate the process of infection [9], while increasing the insect's metabolism causes its undue starvation, in those circumstances the weaker individuals die. The lowering of temperature to 28° brings about a slight delay of red lice occurrence (Fig. 15) due to prolongation of logarithmic phase of rickettsia growth. It has no influence on the infection degree. Similar situation prevails at 24°, with louse reddening being still more delayed.
The environmental temperature of 32° is optimal both for lice and rickettsiae. Its rise to 37° does not essentially accelerate the process of infection [9], while increasing the insect's metabolism causes its undue starvation, in those circumstances the weaker individuals die. The lowering of temperature to 28° brings about a slight delay of red lice occurrence (Fig. 15) due to prolongation of logarithmic phase of rickettsia growth. It has no influence on the infection degree. Similar situation prevails at 24°, with louse reddening being still more delayed.
More noticeable effect is exerted at 22°. At this temperature not only the appearance of red lice is very belated, but the process itself of reddening of infected population is stretched over many days. Much sharper individual differences in the rate of development of infection in particular insects are recorded here. In the first ten days following the inoculation the lice are but weakly infected, this being particularly true of males. LID100 of rickettsiae in those lice ranges from 104 to 106. After the expiration of 10 days 30 p.c. of stomach epithelial cells are found to be infected [9]. The degree of infectivity is equal to 107LID100. About the 15th day 100 p.c. of cells show the presence of rickettsiae. The number of living rickettsiae amounts to 105-6LID100. Identical situation is met at 18°, the whole process being delayed by 3 days in comparison with that at 22°.
V. THE PROCEDURE IN BREEDING OF LICE INFECTED WITH RICKETTSIA PROWAZEKI 1. Breeding of infected lice for culture of strains and production of typhus vaccine
The infected lice should be fed not earlier than 30 min. after rickettsia injection and not later than 6 h. Shorter time would cause the elimination of a great number of rickettsiae with faeces, evacuated intensively by the insect during blood sucking. Later feeding would make the lice excessively famished. Next, the infected individuals, like the healthy ones, are fed once daily for 30 min. They are incubated at 32° [15, 20]. The first survey of a given cage is carried out about the 50th h ] incubation in case of using an adapted strain, and from 3 to 6 h after the last feeding. All red lice are then removed. They may be used for vaccine production, but never for passaging strains, on the ground of their being weakly infected. Next sorting out takes place 72 h after inoculation before feeding and again 3 h after feeding. The liquidation of all infected population occurs when above 80 p-c. of specimens turn red in a cage. When strains of slower rate of development are employed,, the surveys and the liquidation are accordingly delayed. If the process! of infection of a given population is not over within 5 days, the cage and the cloth must be changed on account of larval hatching from eggs laid at that time.
In order to exclude the presence of other kinds of bacteria the faeces must be daily examined on agar plates. If on the first day the lice turn out to be infected with cocci, they must be discarded, whereas the appearance of bacterial flora later on makes the use of these lice possible for vaccine production. In this case a longer storage in 0.5 p.c. phenol before insects preparation is required.
2. Breeding of infected lice in experimental works
In experimental culture of Rickettsia prowazeki, the cages containing infected lice should be examined as early as on the first day after inoculation to establish the number of individuals which had perished due to the damage sustained during the injection. The examination should be performed on every consecutive day and red lice removed as well as those dead of other causes. Both these groups are recorded separately. The experiments are terminated either when about 80-90 p.c. of insects turn red or when, the process of reddening is over, due to the lack of infection in some specimens. It happens when .more diluted suspension or those subjected to rickettsicidal action are used.
VI. THE PROCEDURE IN VACCINE PREPARATION Lice earmarked for vaccine production are put into jars containing 0.5 p.c. phenol solution with the aim of killing rickettsiae, which perish not later than after 96 h [13]. When they are to be used for the preparation of the antigen for Weigl's reaction, they are placed in saline. After soaking of 24-28 h the lice are dissected under a stereoscopic microscope, applying the magnification of 16 times. With the tip of a needle and dissecting scalpel a nick is made between the thorax and abdomen of louse, taking care not to penetrate to the alimentary tract. The thorax is held with the side of the needle and using the tip of the scalpel the abdomen is drawn in a posterior direction. The exposed stomach should be cut off on both sides and removed to a drop of phenol solution or saline. Prepared intestines are collected and placed in tubes containing either 0.5 p.c. phenol solution (vaccine) or saline (antigen).
2. Purification of the vaccine
Prepared louse intestines are homogenized in Weigl's mortar and then suspended in 0,5 p.c. phenol solution or saline. Next, they are centrifuged for 5 minutes at the speed below 1000 r.p.m. The supernatant containing rickettsiae is withdrawn and the residue is washed twice The rickettsial suspension is then subjected to centrifugation during 1 h at the speed of 5000-6000 r.p.m. The supernatant is next rejected and the rickettsiae being in the sediment are suspended in 0.5 p.c. phenol solution in the quantity corresponding to 100 stomachs in 1 ml. After assessing its value in animals, the vaccine is suitably diluted. The proportion of 90 stomachs per one vaccine proposed by Weigl seems too strong if, of course, we are dealing with highly infected lice.
VII. EPIZOOTIES IN LABORATORY COLONIES OF LICE Epizootics among lice are regarded as one of the most difficult problems in Weigl's vaccine production. One group of their causative agents is composed of staphylococci, micrococci, neisseriae and Gram-negative rods such as E. coli, Proteus, Aeromonas and Pseudomonas. The other group is constituted by Rickettsia rocha-limae and Rickettsia quintana s. pediculus weigli.
1. Infections caused by cocci and Gram-negative rods
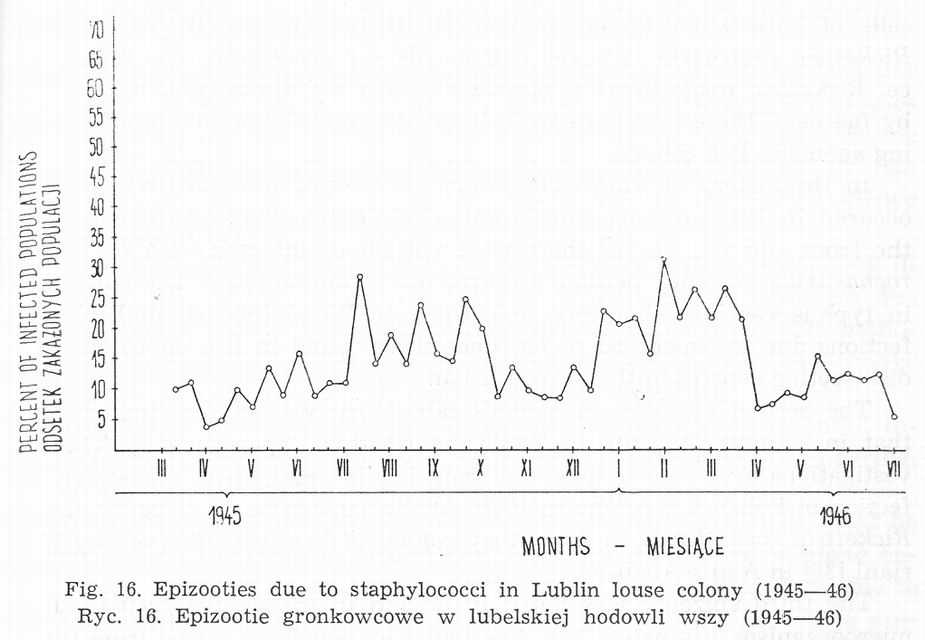
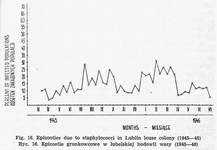 Bacterial infections can originate from human skin, human blood, unsterilized cages and introduction of bacteriae into louse stomach. The course of insect's infection depends on it's route and bacteria species. Staphylococci and micrococci are mostly derived from feeder's skin and are found only on the 'body surface of louse. This is particularly observed when too many individuals are held in a cage. It hinders transpiration and makes the insects wet. Epizootics evoked by cocci are subject to periodical growth and decline in the proportion of infected population [Fig. 16]. Great differences related to individual properties of the feeder's skin are observed, too. Cocci living on the body surface of lice do not cause their death, but introduced into intestines lead to depopulation [27, 30, 31], the course of which is related to staphylococcus strain.
Bacterial infections can originate from human skin, human blood, unsterilized cages and introduction of bacteriae into louse stomach. The course of insect's infection depends on it's route and bacteria species. Staphylococci and micrococci are mostly derived from feeder's skin and are found only on the 'body surface of louse. This is particularly observed when too many individuals are held in a cage. It hinders transpiration and makes the insects wet. Epizootics evoked by cocci are subject to periodical growth and decline in the proportion of infected population [Fig. 16]. Great differences related to individual properties of the feeder's skin are observed, too. Cocci living on the body surface of lice do not cause their death, but introduced into intestines lead to depopulation [27, 30, 31], the course of which is related to staphylococcus strain.
Neisseriae are encountered incomparably less frequently. The infections probably take place only during bacteremia in feeder's blood. After artificial infection, due to neisseriae the lice perish in 40-70 p.c.
Gram-negative rods introduced intraintestinally are fatal to lice and kill them within a few hours [30, 31]. They are also harmful in contact infection, but its course is less rapid and the mortality rate among insects is lower.
2. Infection due to Rickettsia rocha-limae
Nonpathogenic, intracellular rickettsiae were recorded already by Rocha-Lima. The next to observe and to describe them was Weigl [62, 63], who gave the new species the name of Rickettsia rocha-limae. As follows from his investigations Rickettsia rocha-limae is not indentical with extracellular Rickettsia quintana and differs from Rickettsia prowazeki in respect if a whole range of features [62, 63]. First of all, polymorphism of Rickettsia rocha-limae is more pronounced. In smears from infected louse stomach they seem to be agglutinated. Rickettsia rocha-limae stain much more readily -with aniline dyes than Rickettsia prowazeki. In histological picture their intracellular presence is clearly visible. Inside the cells they form irregular masses and contrary to Rickettsia prowazeki they prevent a good vision of single rickettsiae. Rickettsia rocha-limae in spite of abundant multiplication in epithelium cells of louse stomach does not cause its reddening and death. While Rickettsia prowazeki is never responsible for cross-infections among lice, Rickettsia rocha-limae is spread not only by direct contact, but also by faeces of these insects as well. It causes rapid dissemination of infecting agents in lice colony.
In the history of Polish laboratory colonies of lice the first epizooty occurred in 1919. Among insects collected from soldiers returning from the front and war camps there were specimens infected with Rickettsia rocha-limae and they became a source of contamination of lice cultivated in typhus vaccine laboratory. According to Weigl [62, 63] multiple infections due to Rickettsia rocha-limae were found in lice collected from disinsecting centers in the years 1920 and 1921.
The second epizooty in Weigl's laboratory was closely linked with that in Pasteur Institute in Tunis described by Sparrow [54]. The investigations over insects obtained from her brought about a massive infection in Polish lice colony in 1939. At the same time an epizooty due to Rickettsia rocha-limae in laboratory colony of lice was observed by Mariani [35] in Addis-Ababa.
The third epizooty was noted in Poland in the winter 1940/41. This microorganism originated from lice found on people suffering from typhus. This fact had complicated and obscured the problem, as a mixed infection of two rickettsiae: Rickettsia prowazeki and Rickettsia rocha-limae, appeared.
The control of Rickettsia rocha-limae consists in total liquidation of the population affected with epizooty and thorough disinfection of room and incubators. The feeders are of no importance, as Rickettsia rocha-limae is not only nonpathogenic for human beings, but its carriage has not been known [5, 62, 631.
3. Infection due to extracellular rickettsiae
Extracellular rickettsiae living in louse were recorded for the firs1-time in 1916 by Topfer [58]. After his determining of extracellular rickettsia as an etiological factor responsible for trench fever it was called Rickettsia quintana by Schmincke [53] and Rickettsia pediculi by Munk and Rocha-Lima [44]. Mosing [39, 40, 41] described laboratory epidemic due to extracellular rickettsiae, named by him Rickettsia weigli. Herzig-Weiglowa [7, 8] regarded Rickettsia weigli and Rickettsia quintana as being two different pathogenic mutants deriving from non-pathogenic Rickettsia pediculi. Her view was shared by Kostrzewski [10, ll] who was of the opinion that the three above mentioned rickettsiae do not differ in any way. Our own observations made during the war-time work in Weigl's laboratory seem to bear this out.
In lice Rickettsiae quintana multiply in lumen of their stomach exclusively. Other parts of alimentary canal, coeloma, Maipighian tubules and reproductive organs are free of it. Rickettsia quintana settle on the surface of stomach epithelial cells and in histological picture look like a pallisade. In smears from louse intestine Rickettsiae quintana evidence a high degree of polymorphism.
Lice get infected with rickettsiae circulating in blood stream of ill or asymptomatic feeders [59]. No contact or transovarial infections due to Rickettsia quintana have ever being observed. It is totally non-pathogenic for lice [7, 8]. In faeces of those insects Rickettsia quintana survives a long time, which may amount to as much as 18 months [10, 11].
The presence of Rickettsia quintana does not exert any essential influence on parallel infection due to Rickettsia prowazeki, hindering, however, microscopic evaluation of vaccine material.
While passaging Rickettsia prowazeki strains benzylpenicillin must be added to the suspension in case of co-infection of Rickettsia quintana being found during histologic examination of strains employed. It is due to Rickettsia quintana being sensitive to this antibiotic, while Rickettsia prowazeki is not. Rickettsia quintana can be also eliminated by passaging through guinea pigs, mice and yolk sacs of chick embryos, as it cannot be transmitted to them [37, 72].
VIII. HARMFUL EFFECT OF LABORATORY LOUSE FEEDING ON HOST ORGANISM 1. The effect of louse feeding on blood of human feeders
Healthy louse feeder feeds monthly an average of 25.000 specimens from the moment of their hatching until attaining complete sexual maturity. It corresponds to about 75.5 g of blood. Adult and Rickettsia prowazeki infected lice are fed from 4 to 6 times. At the rate of 25.000 individuals monthly the quantity of lost blood amounts to 100-150 g. In spite of that loss of .blood being rather insignificant, some louse feeders display a marked lack of tolerance of louse feeding [6, 56]. This sensitivity is an individual feature, independent of age, sex, constitutional traits and general condition. A sudden diminution of the amount of hemoglobin is noticeable in those people. The louse feeding, however, has no effect on erythropoiesis. According to Tomaszewski [56] about 30 p.c. of human louse feeders show this reaction.
2. Reaction of human skin to louse feeding
Skin reaction at the site of louse feeding can be divided into the early and late one. The early reaction characterized by reddening and itching of the skin appears already during the feeding and maintains from several hours to a few days, this time being dependent on the length of the feeder's status. The longer the feeding, the weaker the reaction.
The late reaction occurs several hours or the next day after louse feeding and is characterized not only toy reddening and itching, but also by swelling and serum exudate. The reaction can persist for as long as one week. Mostly no decline of feeder's sensitivity is observed here. Such people cannot act as louse feeders.
3. Allergic reaction to louse faeces
Allergic reactions due to airborne louse faeces are often encountered in people working with lice. Most frequent here is rhinitis, sometimes associated with conjunctivitis. Cases of bronchial asthma have been also recorded. Individuals with such allergy cannot 'be vaccinated with unpurified Weigl's vaccine. An anaphylactic shock may result then [57].
IX. INFECTIONS OF WORKERS IN LABORATORIES EMPLOYING WEIGL'S METHOD 1. Typhus of vaccinated people
During the last war and immediately after its end many people employed at the production of Weigl's vaccine fell ill with laboratory typhus in spite of prior immunization [42, 43, 68], whereas at the Department of Microbiology in Gdansk concerned with Rickettsia prowazeki culture of lice only single cases of this disease were noted in the years 1946-1973. The latter pertained to people vaccinated about one year before. These facts may be ascribed to lower resistance of organism due to war conditions.
Typhus in vaccinated people has a different course than the classic one, as it was found by Mosing [43]. Its duration does not exceed 1° days, but abortive forms manifesting themselves in a fever of severa hours are also not uncommon. The temperature is high, but not droppi1^ per crisim. Relative bradycardia is one of the most characteristic feature of typhus in vaccinated people. Rash appears rather rarely and has the character of roscola. As a rule, no deaths are recorded among individuals vaccinated with Weigl's vaccine.
First laboratory epidemic of trench fever in Poland was described by Mosing [39, 40, 41] and designated rickettsemia Weigli. Similar infections were later reported by Herzig [7], Petrynski [45], Przybylkiewicz and Kostrzewski [47]. Mosing, Herzig and Petrynski had treated rickettsemia Weigli as a disease entirely different from trench fever, while Kostrzewski [10, 11] saw no differences between war-time trench fever and laboratory rickettsemia Weigli. Our observations made during the II World War seem to confirm this hypothesis. While conducting experiments with Riickettsia quintana isolated from German soldiers, a lot of workers fell ill. The clinical course was much more serious than at the so called rickettsemia Weigli, but lighter than in German soldiers. After some time a great part of staff were affected with this front trench fever. With the lapse of time the course of the illness became milder, with occurrence of abortive and unapparent forms of infection.
Laboratory diseases caused by Rickettsia quintana in wartime concerned not only louse feeders; infections among laundry personal having no direct contact with lice were also recorded. It is due to the enormous role played by louse faeces in spreading this infection.
Laboratory diagnosis of trench fever is based, first of all, on louse feeding test (xenodiagnosis) [37, 38]. It is also the only criterion for assessing the effect of antibiotics on Rickettsia quintana [37]. Serological reactions, microagglutination [36] and complement fixation [46, 73], are also introduced at present.
The carrier state and even late relapses may exist and occur many years after the first manifestation of trench fever [10, 11, 37, 38, 59]. This persistence and periodical recrudescence combine to favour the transmission and spread of the disease, especially among louse feeders in typhus laboratory. Tetracyclines are the best antibiotics for treatment of trench fever [37, 38].
REFERENCES
1. Anderson C.R.: Survival of Rickettsia -prowazeki in different diluents. J. Bacteriol., 1944, 47, 519.
2. Bovarnick M.B., Snyder J.: Respiration of typhus rickettsiae. J. Exp. Med., 1949, 89, &61.
3. Bovarnick M.B., Miller J., Snyder J.: The influence of certain salts, amino acids, sugars and proteins on the stability of rickettsiae. J. Eacteriol., 1950, 59, 509.
4. Chodzko W.: Experience polonaise de la vaccination preventive contre le typhus exanthematique d'apres la methode de Weigl. Bull. Mens. Off. Hyg. Publique, 1933, 25.
5. Durand P. Sparrow H.: Innocuite pour l'homme des rickettsias du type Rickettsia rocha-limae. Arch. Inst. Pasteur (Tunis), 1939, 28, 74.
6. Finkel A.: Obraz cytologiczny krwi u ludzi karmiacych wszy. Med. Dosw. Spol., 193.2, 18, 257.
7. Herzig A.: Eine neue Rickettsia-Spezies der Laus, der Erreger einer spontan aufgetretenen epidemischen Erkrankung des Menschen. Zentralbl. Bakteriol. (Orig.), 1939 143, 299.
8. Herzig-Weiglowa A.: Badania nad drobnoustrojem Rickettsia. pediculi. Rozprawy Wydz. Lek. PAU, Krakow, 1947, 8/1, 1.
9. Herzig-Weiglowa A., Krynski S.: Influence de la temperature sur Revolution de l'infection des pous. Arch. Inst. Pasteur (Tunis), 1959, 36, 435.
0. Kostrzewski J.: The epidemiology of trench fever. Bull. Int. Acad. Pol. Sci. Let. (Med.), 1949, 70, 233.
11. Kostrzewski J.: Epidemiologia goraczki. okopowej. Med. Dosw. Mikrobiol, 1950, 2, 287.
12. Krynski S.; Investigations on the toxic action of Rickettsia prowazeki on lice and white mice. Bull. Inst. Mar. Trop. Med. (Gdansk), 1948, 3, 33.
13. Krynski S,, Czuczwar S.: Badania nad dzialaniem fenolu na Rickettsia prowazeki. Przegl. Epidemid., 1948, 2, 2
14. Krynski S.: Badania nad toksycznym dzialaniem zarazka duru osutkowego; Rickettsia prcwazeki. Przegl. Epidemiol., 1,948, 2, 50.
15. Krynski S.: Zasady hodowli wszy sztucznie zakazonych zarazkiem duru osutkowego metoda Weigla. Now. Lek., 1948, 25, 267.
16. Krynski S., Woyciechowska S.: Influence of various factors upon .virulence and vitality of Rickettsia prowazeki in laboratory breeding by Weigl's method. Bull. Inst. Mar. Trop. Med. (Gdansk), 1949, 2, 49.
17. Krynski S.: Forms of Rickettsia prowazeki infection in lice artificially infected by Weigl's method. Bull. Inst. Mar. Trop. Med. (Gdansk), 1949, 2, 231.
18. Krynski S., Woyciechowska S.: Badania nad dzialaniem siarczanu miedzi na Rickettsia prowazeki w hodowli laboratoryjnej metoda Weigla. Przegl. Epidemiol., 1949, 3, 94.
19. Krynski S.: Zasady przygotowania zawiesin do sztucznego zakazania wszy metoda Weigla. Przegl. Epidemiol., 1949, 3, 333.
20. Krynski S., Radkowiak J.D.: Principles of cultivation of Rickettsia prowazeki by Weigl's method. Bull. Stat. Inst. Mar. Trop. Med. (Gdansk), 1952, 4, 213 (in Polish, Russian and English).
21. Krynski S., Becla E.: Influence of milk on viability and infectivity of Rickettsia prowazeki. Bull. Stat. Inst. Mar. Trop. Med. (Gdansk), 1955, 6, 193 (in Polish, Russian and English).
22. Krynski S., Becla E.: Influence de la temperature sur l'activite toxique de Rickettsia prowazeki chez Ie pou. Arch. Inst. Pasteur (Tunis), 1960, 37, 3. -
23. Krynski S., Becla E.: Infection a P.pseudo-tuberculosis chez le pou. Ann. Inst. Pasteur (Paris), 1963, 104, 133.
24. Krynski S., Becla E.: Infection of body louse with Pasteurella pseudotuberculosis after intrarectal inoculation. J. Infect. Dis., 1964, 114, 379.
25. Krynski S. Becla E.; The action of oxytetracycline on Rickettsia prowazeki in lice inoculated by the Weigl method. Chemotherapy, 1964, 8, 265.
26. Krynski S., Becla E.: Courbe de croissance de Rickettsia prowazeki dans l'intestin du pou infecte selon la methode de Weigl. Arch. Inst. Pasteur (Tunis), 19.6,6, 43, 365.
27. Krynski S., Becla E.: Staphylococcal infections in lice. Postepy Mikrobiol., 1966, 5, 327 in English).
28. Krynski S., Mollaret H.H., Becla E.: L'inoculation le Yersinia enterocolitica par voie intrarectal chez le pou. Ann. Inst. Pasteur (Paris 1966, 110, 779).
29. Krynski S.: Rudolf Weigl (1883-1957). Exp. Med. Microbbiol., 1967, 19, 233.
30. Krynski S.: Bacterial infections in lice injected by Weigl method. Wiad. Parazytol., 1967, 13, 615 (in English).
31. Krynski S.: Influence of individual features and environmental temperature on the development of infection and intoxication in lice. The Control of Lice and Louse-borne Diseases. Pan American Health Organization, Scientific Publication, 1973, 23, 162.
32. Machel M., Becla E.: A new technique for parenteral inoculation of the tick Ornithodoros moubata (Murray). Z. Angew. Zool., 197,0, 57, '257.
33. Mariani G.: Sulla vaccinazione controle rickettsiosi humane. Ann. Ig. (Roma), 1938, 48, 3'72.
34. Mariani G.: Vaccinazioni contro il tifo esantematico eseguite nel 1938 sullaltipiano etiopico con il vaccino Weigl. Ann. Ig. (Roma), 1939, 49, 316.
35. Mariani G.: Caractteristico del ceppo etiopico de Rickettsia rocha-limae. Ann. Ig. (Roma), 1940, 50, 59.
36. Meyers W.F., Wisseman Ch.L.Jr.: Serological studies of trench fever employing a microagglutination procedure. The Control of Lice and Louseborne Diseases. Pan American Health Organization, Scientific Publication, 1973, 263, 79.
37. Mohr W., Weyer F.: Neuere Beobachtungen ueber Wolhynisches Fieber, insbesondere ueber das Auftreten von Spatruckfallen. Dtsch. Arch. Klin. Med., 1964, 2'09, 392.
38. Mohr W., Weyer F.: Late relapses of trench fever. Ger. Med. Mon., 19i84, 9, 145
39. Mosing H.: Rickettsemia Weigli. Pol. Gaz. Lek., 1935, 14, 8.
40. Mosing H.: Une nouvelle infection a Rickettsia: Rickettsia weigli nov. sp. Arch. Inst. Pasteur (Tunis), 1936, 25. 373.
41. Mosing H.: Badania nad epidemia wywołaną przez nowy typ rickettsji Rickettsia weigli. Med. Dosw. Spot., 193.6, 21, ,218.
42. Mosing H.: Methods of evaluation of typhus vaccine potency. Tex. Rep. Biol. Med., 1947, 5, 173.
43. Mosing H.: Tyfus plamisty u osób szczepionych. Rozprawy Wydz. Lek. PAU. Krakow, 194'8, 8/4, 1.
44. Munk F., da Rocha-Lima H.: Klinik und Aetiologie des sogen. "Wolhynischen Fiebers" (Werner-Hissche Krankheit). II. Ergebnis der atiologischen Untersuchungen und deren Beziehungen zur Fleckfieberforschung. Munch. Med. Wochenschr., 1917, 64, 1422.
45. Petrynski B.: W sprawie nowej endemii w Zakładzie Biologii U.J.K. Sprawozdania PAU, 1948, 49, 20'2.
46. Pschetnichnov R.A., Plaksina A.N.: Diagnostica wolynskogo rikketsioza po reakcii swiazywanija komplementa. Laboratornoje Dielo, 1965, 12, 732.
47. Przybylkiewicz Z., Kostrzewski J.: Zaraza pracowniana goraczki okopowej. Przegl. Lek., 1948. 4, 782.
48. Da Rocha-Lima H.: Untersuchungen ueber Fleckfieber. Munch. Med. Wochenschr., 1916, ,63, 1381
49. Radkowiak J.D.: Technika zakazania wszy metoda Weigla. Przegl. Epidemiol., 1949, 3, 189.
50. Radlo P.: Observations sur la vaccination centre le typhus exanthematique par vaccin de Weigl. Arch. Inst. Pasteur (Tunis), 1937, 26, 667.
51. Rutten J.: La mortalite des missionaires avant et apres l'emploi du |vaccin de Weigl. Dossiers de la Commission Synodale a Peking, 19'3'6, 183.
52. Rutten J.: Dernier resultats des vaccinations contre le typhus. Dossiers de la Commissions Synodale a Peking, 1943, 79.
53. Schmincke R.A.: Histopatologischer Befundsin Roseolen der Haut bei Wolhynischem Fieber. Munch. Med. Woichenschr., 1917, 64, 61.
54. Sparrow H.: Infection spontanee des pous d'elevage par une rickettsia du type Rickettsia rocha-limae. Arch. Inst. Pasteur (Tunis), 1939, 28, 64.
55. Starzyk J.: Vitalite, virulence et pouvoir immunisant de Rickettsia prowazeki conserves en dehors de l'organisme du pou. Arch. Inst. Pasteur (Tunis), 1938, 27, 263.
56. Tomaszewski L.: Wplyw karmienia wszy na sklad krwi obwodowej karmicieli, ze specjalnym uwzglednieniem retikulocytow. Sprawozdania PAU, 1948, 49, 53.
57. Tomaszewski L.: Ogolne odczyny poszczepienne przy uodparnianiu szczepionka Weigla w Lubelskim Zakladzie Wyrobow Szczepionki przeciw Durowi Plamistemu im. R. Weigla. Sprawozdania PAU,1948, 49, 205.
58. Topfer H.: Zur Ursache und Uebertragung des Wolhynischen Fiebers. Munch. Med. Wochenschr. 1916, 63, 1495.
59. Vinson J.W.: Geographic distribution of trench fever. The Control of Lice and Louse-borne Diseases, Pan American Health Organization, Scientific Publication., 1973, 2,63. 76.
60. Weigl R.: Untersuchungen und Experimente an Fleckfieberlausen. Die Technik der Rickettsia-Forschung. Beitr. Klin. Infectionskrank., 1919, 8, 353.
61. Weigl R.: Badania nad Rickettsia prowazeki. Przegl. Epidemiol., 1920, 1,4, -
62. Weigl R.: Dalsze badania nad Rickettsja. Rickettsia rocha-limae (nov. spec.), Przegl. Epidemiol., 1921, 1, 375.
63. Weigl R.: Further studies on Rickettsia rocha-limae (nov. spec.), J. Trop. Med. Hyg., 1924, 27, 14.
64. Weigl R.: Ueber active Fleckfieberimmunitat. Med. Klin., 1924, 20, 105'6. -
65. Weigl R.: Die Methoden der activen Fleckfieber-Immunisierung. Bull. Int. Acad. Pol. Sei. Let. (Med.), 1930, 25.
66. Weigl R.. Sposoby czynnego uodparniania przeciw durowi osutkowemu. Rozprawy Wydz. Lek. PAU, Krakow, 1931, 1/1, 1.
67. Weigl R.: La serovaccination de l'homme contre le typhus. Arch. Inst. Pasteur (Tunis), 1937, 26, 665.
68. Weigl R., Mosing H.: Uodpornienie czlowieka przeciw durowi osutkowemu według kombinowanej metody Weigla. Sprawozdania PAU, 194'6, 47, 223.
69. Weigl R.: Immunization against typhus fever in Poland during World War II. Tex. Rep. Biol. Med., 1947, 5, 177.
70. Weigl R.: Metody walki z durem osutkowym. Przegl. Epidemiol., 1948, 2, 3.
71. Weyer F.: Die kunstliche Infection von Zecken mit Rickettsien und anderen Krankheitserregern. Zentralb. Bakteriol. (Orig.), 1943, 152, 449.
72. Weyer F.: Epidemiologie des rickettsioses et en particulier de la fievre des tranchees. Donnees nouvelles. BuL Soc. Pathol. Exot., 1963, 56, 590.
73. Weyer F., Vinson J.W., Mannweiler E., Mohr W.: Serologische Untersuchungen bei Wolhynischem Fieber. Z. Tropenmed. Parasitol., 1972, 23, 187.
74. Wohlrab R.: Fleck-typhusbekampfung in Generalgouvernement. Munch. Med. Wochenschr., 1942, 89. 483.
75. Woyciechowska S.: Oczyszczanie szczepow Rickettsji prowazeki z rickettsji zewnatrzkomorkowych sposobem zakazania wszy silnie rozcienczonymi zawiesinami. Sprawozdania PAU, 1947, 43, 424.
76. Woyciechowska S.: 0 barwieniu rickettsji. Przegl. Epidemiol., 1949, 3, 3o4.
METODA WEIGLA DOODBYTNICZEGO ZAKAZANIA WSZY W PRODUKCJI SZCZEPIONKI PRZECIW DUROWI PLAMISTEMU I PRACACH DOSWIADCZALNYCH Z RICKETTSIA PROWAZEKI Streszczenie
W polskich laboratoriach hodujących wszy jest używany szczep Weigla, stanowiący hybrydę rasy kaukaskiej i afrykańskiej. Wskutek wieloletniej wsobnej hodowli w warunkach laboratoryjnych, biologia tych wszy uległa wielu zmianom, szczególnie dotyczy to granic temperatur, w jakich ten owad się rozwija i rozmnaża. Wszy są trzymane w specjalnych klatkach. W Polsce najczęściej używany jest typ Sikory. Karmi się je raz dziennie przez 30 min. na ludziach. Owady dojrzale w 10- 12 dniu życia zakaża się doodbytniczo specjalną mikrokapilarą po uprzednim unieruchomieniu w imadełkach. Płyn jest wprowadzany pod ciśnieniem przy zastosowaniu elektromagnetycznego przerywacza. Wszy zakażone karmi się przez 3-6 dni. Na przebieg procesu zakażenia mają wpływ szczep i liczba jego pasaży przez wszy, wielkość dawki zakażającej i temperatura otoczenia. Zakażone owady staja się czerwone na skutek przenikania hemoglobiny, a nawet całych czerwonych krwinek z przewodu pokarmowego do jamy ciała. Wysoko zakażona wesz zawiera 107 LID100 R. prowazeki. Osobniki przeznaczone na szczepionkę umieszcza się w 0,5 procentowym fenolu, a po wypreparowaniu jelit rozciera się je, a następnie zawiesinę wiruje celem oddzielenia rikettsji od tkanek owada.
Wśród wszy bywają epizootie wywoływane przez gronkowce, neisseriae, Gram-ujemne pałeczki, oraz 2 gatunki rikettsji: R. rocha-limae i R. quintana. U ludzi karmiących wszy może wystąpić spadek ilości hemoglobiny, odczyny skórne oraz alergiczny nieżyt nosa, a nawet dychawica oskrzelowa. Pracownicy mogą równiez zapadać na poronny dur plamisty i gorączkę okopową.
Adres autorow: Zakład Mikrobiologii AMG, ul. Hibnera 38, 80-227 Gdańsk-Wrzeszcz.
Copyright 1974 Stefan Kryński
All rights reserved
Wszystkie prawa zastrzeżone.